Genetic course lecture (2) Extension to Mendelian analysis


























34 Slides2.74 MB
Genetic course lecture (2) Extension to Mendelian analysis (single gene inherence, dominance , lethal alleles , multiple alleles, pleiotropy)
Extensions to Mendelian analysis: Beginning with the first decade of the twentieth century, geneticists subjected many kinds of plants and animals to controlled breeding tests, using Mendel’s 3: 1 phenotypic ratio as a guideline. If the traits under analysis behaved as predicted by Mendel’s laws, then they were assumed to be determined by a single gene with alternative dominant and recessive alleles. Many traits, however, did not behave in this way. For some, no definitive dominance and recessiveness could be observed, or more than two alleles could be found in a particular cross. Other traits turned out to be multifactorial , that is, determined by two or more genes, or by the interaction of genes with the environment. The seed coat color of lentils is a
Extensions to Mendelian analysis include: Term Meaning 1- Incomplete dominance : Pattern of heredity in which one allele is not completely dominant over another 2- Codominance : Pattern of heredity in which both alleles are simultaneously expressed in the heterozygote 3- Multiple alleles : A gene that is controlled by more than two alleles 4- Pleiotropy : When one gene affects multiple characteristics 5- Lethal allele : Allele that results in the death of an individual
Extensions to Mendel for Single-Gene Inheritance we learned that the principle of segregation and the principle of independent assortment allow us to predict the outcomes of genetic crosses. Here, we examine several additional factors acting at individual loci that can alter the phenotypic ratios predicted by Mendel’s principles. One of Mendel’s important contributions to the study of heredity is the concept of dominance—the idea that an individual organism possesses two different alleles for a characteristic but the trait encoded by only one of the alleles is observed in the phenotype. With dominance, the heterozygote possesses the same phenotype as one of the homozygotes. Mendel observed dominance in all of the traits that he chose to study extensively, but he was aware that not all characteristics exhibit dominance. He conducted some crosses concerning the length of time that pea plants take to flower. dominance is frequently “in the eye of the beholder,” meaning that the classification of dominance depends on the level at which the phenotype is examined.
Types of Dominance 1- complete dominance 2- incomplete dominance 3- codominance 4- over dominance
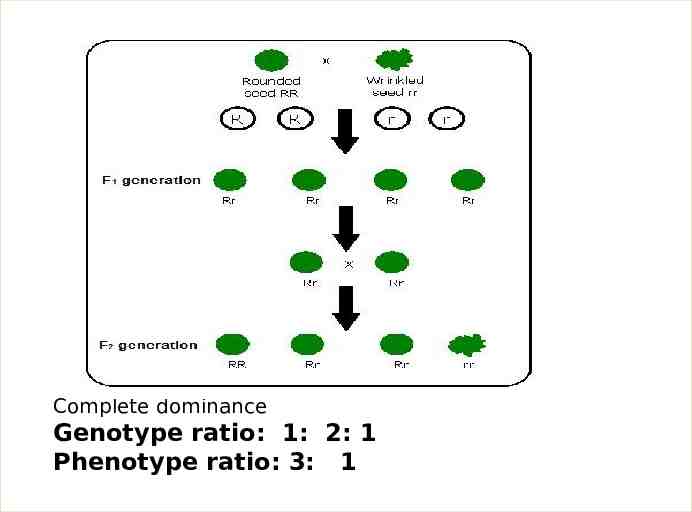
1- Complete dominance In the complete dominance, the effect of one allele in a heterozygous genotype completely masks the effect of the other. The allele that masks the other is said to be dominant to the latter, and the allele that is masked is said to be recessive to the former. Complete dominance, therefore, means that the phenotype of the heterozygote is indistinguishable from that of the dominant homozygote. A classic example of dominance is the inheritance of seed shape (pea shape) in peas. Peas may be round (associated with allele R) or wrinkled (associated with allele r). In this case, three combinations of alleles (genotypes) are possible: RR and rr are homozygous and Rr is heterozygous. The RR individuals have round peas and the rr individuals have wrinkled peas. In Rr individuals the R allele masks the presence of the r allele, so these individuals also have round peas. Thus, allele R is completely dominant to allele r, and allele r is recessive to allele
Complete dominance Genotype ratio: 1: 2: 1 Phenotype ratio: 3: 1
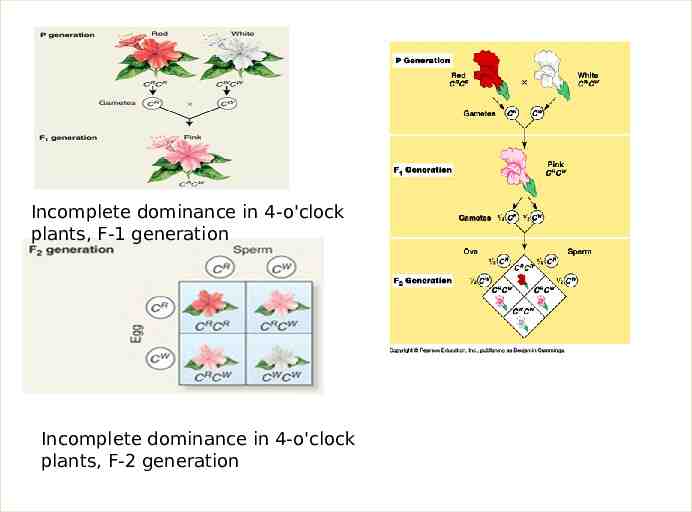
2- Incomplete dominance (also called partial dominance, semi-dominance or intermediate inheritance) occurs when the phenotype of the heterozygous genotype is distinct from and often intermediate to the phenotypes of the homozygous genotypes. For example, the snapdragon flower color is homozygous for either red or white. When the red homozygous flower is paired with the white homozygous flower, the result yields a pink snapdragon flower. The pink snapdragon is the result of incomplete dominance. A similar type of incomplete dominance is found in the four o'clock plant wherein pink color is produced when true-bred parents of white and red flowers are crossed. In quantitative genetics, where phenotypes are measured and treated numerically, if a heterozygote's phenotype is exactly between (numerically) that of the two homozygotes, the phenotype is said to exhibit no dominance at all, i.e. dominance exists only when the heterozygote's phenotype measure lies closer to one homozygote than the other. When plants of the F1 generation are self-pollinated,
Incomplete dominance in 4-o'clock plants, F-1 generation Incomplete dominance in 4-o'clock plants, F-2 generation
In such a case, F2 phenotypic ratio and genotypic ratio are the same, as follows : F2 phenotypic ratio 1 Red : 2 Pink : 1 White F2 genotypic ratio 1 RR : 2 Rr : 1 rr The modern biochemical explanation for this type of incomplete dominance is that each allele of the gene under analysis specific es an alternative form of a protein molecule with an enzymatic role in pigment production. If the “white” allele does not give rise to a functional enzyme, no pigment appears. Thus, in snapdragons and fouro’clocks, two “red” alleles per cell produce a double dose of a red-producing enzyme, which generates enough pigment to make the flowers look fully red. In the heterozygote, one copy of the “red” allele per cell results in only enough pigment to make the flowers look pink. In the homozygote for the “white” allele, where there is no functional enzyme and thus no red pigment, the flowers appear white.
Another example of an intermediate expression may be the pitch of human male voices. The lowest and highest pitches apparently are found in men who are homozygous for this trait (AA and aa), while the intermediate range baritones are heterozygous (Aa). The child-killer disease known as Tay-Sachs* is also characterized by incomplete dominance. *Heterozygous individuals are genetically programmed to produce only 40-60% of the normal amount of an enzyme that prevents the disease. Tay–Sachs disease (also known as GM2 gangliosidosis or hexosaminidase A deficiency ) is a rare autosomal recessive genetic disorder. In its most common variant (known as infantile Tay–Sachs disease), it causes a progressive deterioration of nerve cells and of mental and physical abilities that begins around six months of age and usually results in death by the age of four. The disease occurs when harmful quantities of cell membrane components known as gangliosides accumulate in the brain's nerve cells, eventually leading to the premature death of the cells. A ganglioside is a form of sphingolipid, which makes Tay–Sachs disease a member of the sphingolipidoses. There is
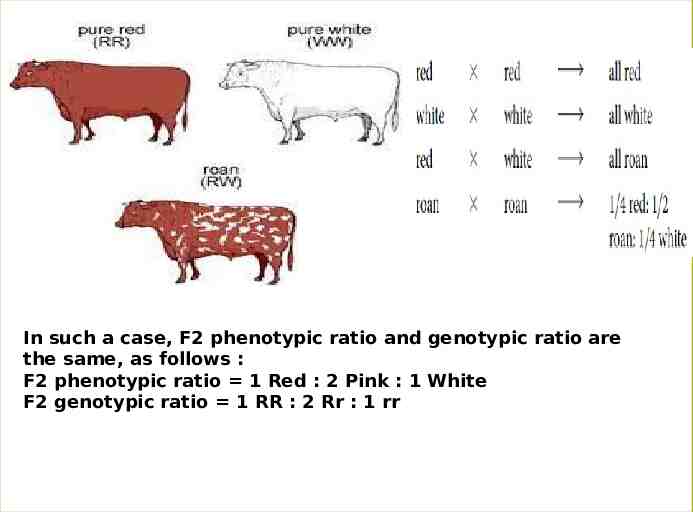
3-Codominance: Codominance Another type of interaction between alleles is codominance, in which the phenotype of the heterozygote is not intermediate between the phenotypes of the homozygotes; rather, the heterozygote simultaneously expresses the phenotypes of both homozygotes. That means . both alleles of a gene in a heterozygote lack the dominant and recessive relationship, i.e., each allele is capable of some degree of phenotypic expression. In a sense, codominance is no dominance Hence, As for incomplete dominance, each of the three genotypes produces its own unique phenotype the phenotypic and genotypic ratios coincide. Examples. 1. The coat color of the Shorthorn breed of cattle represents a classical example of codominance. When a cattle of red coat (CRCR) is crossed with the cattle of white coat (CWCW), the F1 heterozygote or hybrid is found to possess roan coat (CRCW) . In roan coat the red and white hairs occur in definite patches but no hair has intermediate color of red and white.
In such a case, F2 phenotypic ratio and genotypic ratio are the same, as follows : F2 phenotypic ratio 1 Red : 2 Pink : 1 White F2 genotypic ratio 1 RR : 2 Rr : 1 rr
2- The MN locus encodes one of the types of antigens on red blood cells. Unlike antigens foreign to the ABO and Rh blood groups (which also encode red-blood-cell antigens), foreign MN antigens do not elicit a strong immunological reaction; therefore, the MN blood types are not routinely considered in blood transfusions. At the MN locus, there are two alleles: the LM allele, which encodes the M antigen; and the LN allele, which encodes the N antigen. Homozygotes with genotype LMLM express the M antigen on their red blood cells and have the M blood type. Homozygotes with genotype LNLN express the N antigen and have the N blood type. Heterozygotes with genotype L M LN exhibit codominance and express both the M and the N antigens; they have blood-type MN. Genotype Reaction with antisera Anti–M Anti–N Blood group LMLM M LMLN MN LNLN NN
3- in the ABO blood group system, chemical modifications to a glycoprotein (the H antigen) on the surfaces of blood cells are controlled by three alleles, two of which are co-dominant to each other (IA, IB) and dominant over the recessive i at the ABO locus. The IA and IB alleles produce different modifications. The enzyme coded for by IA adds an Nacetylgalactosamine to the membrane-bound H antigen. The IB enzyme adds a galactose. The i allele produces no modification. Thus IA and IB alleles are each dominant to i (IAIA and IAi individuals both have type A blood, and IBIB and IBi individuals both have type B blood, but IAIB individuals have both modifications on their blood cells and thus have type AB blood, so the IA and IB alleles are said to be co-dominant).
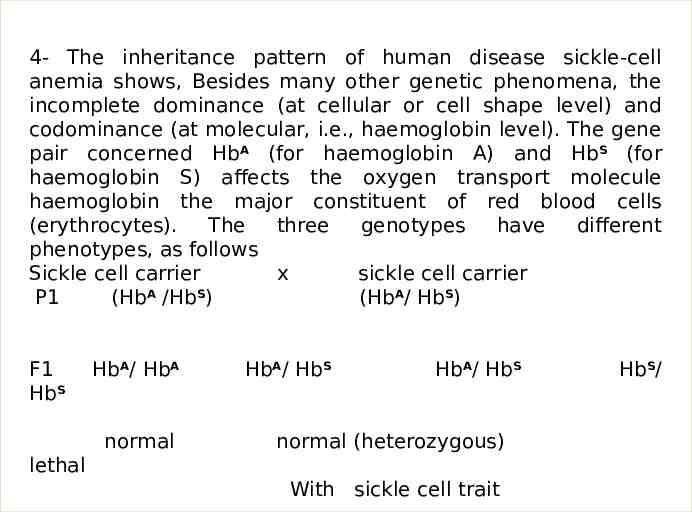
4- The inheritance pattern of human disease sickle-cell anemia shows, Besides many other genetic phenomena, the incomplete dominance (at cellular or cell shape level) and codominance (at molecular, i.e., haemoglobin level). The gene pair concerned HbA (for haemoglobin A) and HbS (for haemoglobin S) affects the oxygen transport molecule haemoglobin the major constituent of red blood cells (erythrocytes). The three genotypes have different phenotypes, as follows Sickle cell carrier x sickle cell carrier P1 (HbA /HbS) (HbA/ HbS) F1 HbS HbA/ HbA normal HbA/ HbS HbA/ HbS normal (heterozygous) lethal With sickle cell trait HbS/
HbA HbA : Normal. Red blood cells never sickled; they contain one type of haemoglobin, i.e haemoglobin A., HbS HbS : Severe, often fatal anemia. Red blood cells sickled-shaped; contain one type of haemoglobin, i.e., haemoglobin S HbA HbS : No anemia. Red blood cells sickle-shaped only under abnormally low oxygen concentration contain both types of haemoglobins, i.e., haemoglobin A and haemoglobin S. Thus, in regard to anemia the HbA HbS HbS allele is dominant. In regard to blood cell shape there is incomplete dominance. And lastly, in regard to haemoglobin, there is codominance. The HbS allele in homozygous condition (HbS HbS ) acts as a lethal gene, i.e., it causes the death of its bearer; the homozygotes dies of fatal anemia before they attain sexual maturity. A marriage between two carriers (i.e., heterozygotes possessing a deleterious recessive allele hidden from phenotypic expression by the dominant normal allele) results in carriers and disease free children in the ratio of 3 : 1, that changes, later on, into the ratio of 2 : 1 due to the
Over dominance In some instances, offspring can demonstrate a phenotype that is outside the range defined by both parents. In particular, the phenomenon known as over dominance occurs when a heterozygote has a more extreme phenotype than that of either of its parents. Indeed, in a few examples, a trait that shows over dominance sometimes confers a survival advantage in the heterozygote
Differences between dominance, codominance Type of Dominance Definition Dominance the incomplete dominance, and Phenotype of the heterozygote is the same as phenotype of one of the homozygotes. Incomplete dominance intermediate (falls Phenotype of the heterozygote is within the range) between the phenotypes of the two homozygotes. over Codominance dominance phenotypes phenotype of heterozygote produce more Phenotype of the heterozygote includes the extreme or better adapted than that of the homozygote of both homozygotes.
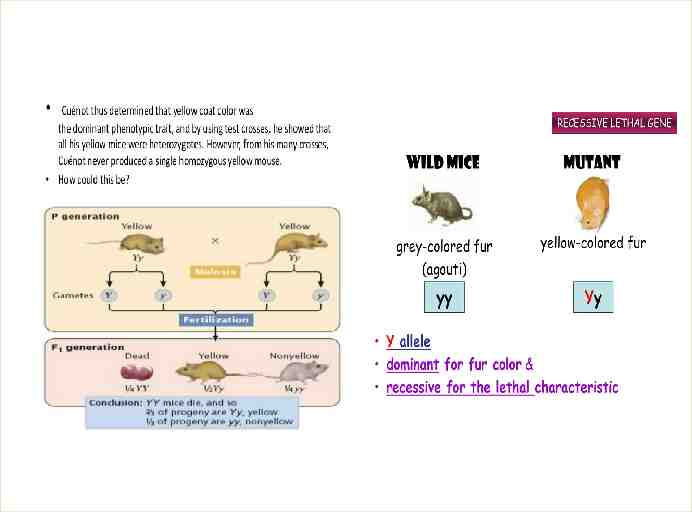
lethal alleles Lethal alleles (also referred to as lethal genes or lethals) are alleles that cause the death of the organism that carries them. They are usually a result of mutations in genes that are essential for growth or development. Lethal alleles can cause death of an organism prenatally or any time after birth, and this would alter the 1:2:1 genotypic ratios and the 3:1 phenotypic ratios predicted for the F 2 generation Lethal alleles were first discovered by Lucien Cuénot in 1905 while studying the inheritance of coat colour in mice. the coat colors of mice are typical Mendelian traits, which have various colors such as white, black, yellow and agouti. The inheritance of mouse coat color is usually stated as an example only in teaching the knowledge of recessive lethal alleles. The agouti gene in mice is largely responsible for determining coat colour. The wild-type allele produces a blend of yellow and black pigmentation in each hair of the mouse. This yellow and black blend may be referred to as
One of the mutant alleles of the agouti gene results in mice with a much lighter, yellowish colour. When these yellow mice were crossed with homozygous wild-type mice, a 1:1 ratio of yellow and dark grey offspring were obtained. This indicated that the yellow mutation is dominant, and all the parental yellow mice were heterozygotes for the mutant allele. By mating two yellow mice, Cuénot expected to observe a usual 1:2:1 Mendelian ratio of homozygous agouti to heterozygous yellow to homozygous yellow. Instead, he always observed a 1:2 ratio of agouti to yellow mice. He was unable to produce any mice that were homozygous for the yellow agouti allele.
Types of lethal allele Recessive lethals: A pair of identical alleles that are both present in an organism that ultimately results in death of that organism are referred to as recessive lethal alleles. example of a recessive lethal allele occurs in the Manx cat. Manx cats possess a heterozygous mutation resulting in a shortened or missing tail. Dominant lethals Alleles that need only be present in one copy in an organism to be fatal are referred to as dominant lethal alleles. These alleles are not commonly found in populations because they usually result in the death of an organism before it can transmit its lethal allele on to its offspring. An example in humans of a dominant lethal allele is Huntington's disease, a rare neurodegenerative disorder that ultimately results in death. Conditional lethals Alleles that will only be fatal in response to some environmental factor are referred to as conditional lethals. One example of a conditional lethal is favism, a sex-linked inherited condition that causes the carrier to develop hemolytic anemia when they eat fava beans
Multiple Alleles: Most of the genetic systems that we have examined so far consist of two alleles. In Mendel’s peas, for instance, one allele encoded round seeds and another encoded wrinkled seeds; in cats, one allele produced a black coat and another produced a gray coat. For some loci, more than two alleles are present within a group of organisms—the locus has multiple alleles. (Multiple alleles may also be referred to as an allelic series.) is a type of non-Mendelian inheritance pattern . Although there may be more than two alleles present within a group of organisms, the genotype of each individual diploid organism still consists of only two alleles. The inheritance of characteristics encoded by multiple alleles is no different from the inheritance of characteristics encoded by two alleles, except that a greater variety of genotypes and phenotypes are possible. Most of the time, when multiple alleles come into play for a trait, there is a mix of types of dominance patterns that occur. Sometimes, one of the alleles is completely recessive to the others and will be masked by any of those that are dominant to it. Other alleles may be co-dominant together and show their traits equally in the phenotype of the individual. There are also some cases where some alleles exhibit
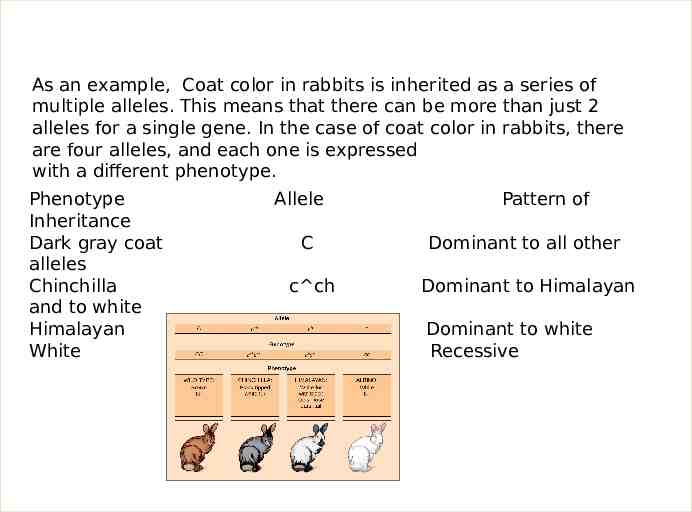
As an example, Coat color in rabbits is inherited as a series of multiple alleles. This means that there can be more than just 2 alleles for a single gene. In the case of coat color in rabbits, there are four alleles, and each one is expressed with a different phenotype. Phenotype Inheritance Dark gray coat alleles Chinchilla and to white Himalayan White Allele Pattern of C Dominant to all other c ch Dominant to Himalayan c h c Dominant to white Recessive
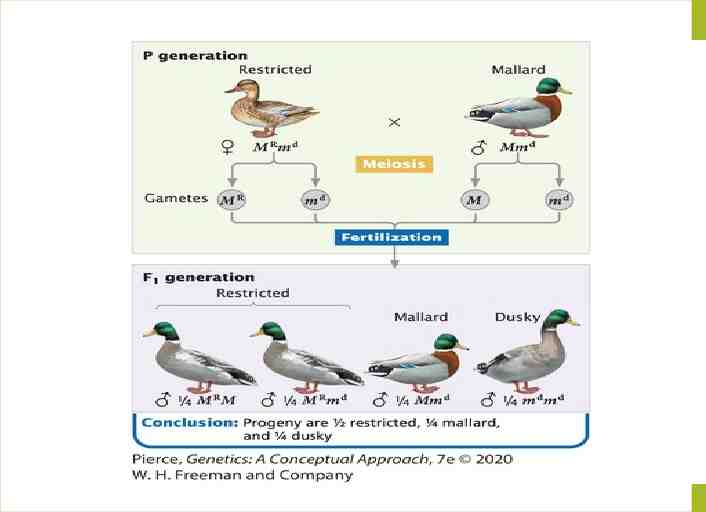
Duck-feather patterns An example of multiple alleles is at a locus that determines the feather pattern of mallard ducks. One allele, M, produces the wild-type mallard pattern. A second allele, MR, produces a different pattern called restricted, and a third allele, md, produces a pattern termed dusky. In this allelic series, restricted is dominant over mallard and dusky, and mallard is dominant over dusky: MR M md. The six genotypes possible with these three alleles and their resulting phenotypes are: Genotype Phenotype MRMR restricted MRM restricted MRmd restricted MM mallard Mmd mallard mdmd dusky
The ABO blood group Another multiple-allele system is at the locus for the ABO blood group. This locus determines your ABO blood type and, like the MN locus, encodes antigens on red blood cells. The three common alleles for the ABO blood group locus are: IA, which encodes the A antigen; IB, which encodes the B antigen; and i, which encodes no antigen (O). We can represent the dominance relations among the ABO alleles as follows: IA i, IB i, IA IB. Both the IA and the IB alleles are dominant over i and are codominant with each other; the AB phenotype is due to the presence of an I A allele and an IB allele, which results in the production of A and B antigens on red blood cells. A person with genotype ii produces neither antigen and has blood type O. The six common genotypes at this locus and their phenotypes . Antibodies are produced against any foreign antigens . For instance, a person having blood-type A produces anti-B antibodies, because the B antigen is foreign. A person having blood-type B produces anti-A antibodies, and someone having blood-type AB produces neither anti- A nor anti-B antibodies, because neither A nor B antigen is foreign. A person having blood-type O possesses no A or B
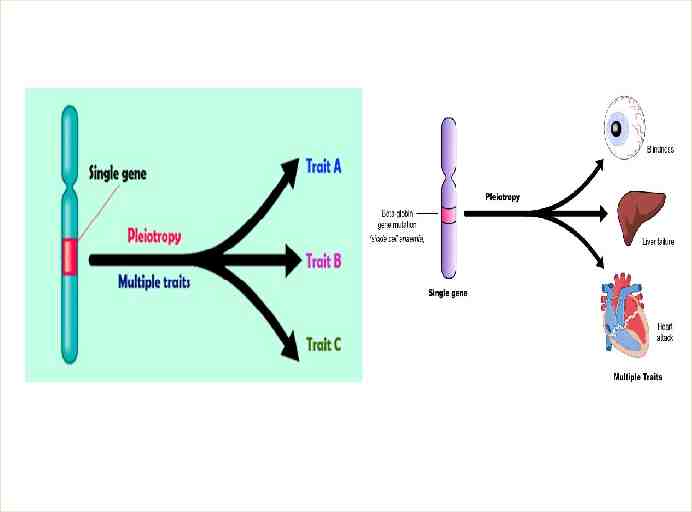
Extensions to Mendel for multifactorial Inheritance One gene may contribute to several characteristics Mendel derived his laws from studies in which one gene determined one trait; but, always the careful observer, he himself noted possible departures. In listing the traits selected for his pea experiments, he remarked that specific seed coat colors are always associated with specific flower colors. The phenomenon of a single gene determining a number of distinct and seemingly unrelated characteristics is known as pleiotropy . Because geneticists now know that each gene determines a specific protein and that each protein can have a cascade of effects on an organism, we can understand how pleiotropy arises. Among the aboriginal Maori people of New Zealand, for example, many of the men develop respiratory problems and are also sterile. Researchers have found that the fault lies with the recessive allele of a single gene. The gene’s normal dominant allele specific a protein necessary for the action of cilia and flagella, both of which are hairlike structures extending from the surfaces of some cells. In men who are homozygous for the recessive allele, cilia that normally clear the airways fail to work effectively, and flagella that normally propel sperm fail to do their job. Thus, one gene determines
That means the PLEIOTROPY Not only is it true that a phenotype can be influenced by many genes, but it is also true that a gene can influence many phenotypes. When a gene affects many aspects of the phenotype, it is said to be pleiotropic, from the Greek words for “to take many turns.” Another example of pleiotropy : The gene for phenylketonuria in humans. The primary effect of recessive mutations in this gene is to cause toxic substances to accumulate in the brain, leading to mental impairment. However, these mutations also interfere with the synthesis of melanin pigment, lightening the color of the hair; therefore, individuals with PKU frequently have light brown or blond hair. Biochemical tests also reveal that the blood and urine of PKU patients contain compounds that are rare or absent in normal individuals. This array of phenotypic effects is typical of most genes and results from interconnections between the biochemical and cellular pathways that the genes control.
Another example of pleiotropy comes from the study of mutations affecting the formation of bristles in Drosophila. Wild-type flies have long, smoothly curved bristles on the head and thorax. Flies homozygous for the singed bristle mutation have short, twisted bristles on these body parts—as if they had been scorched. Thus, the wild-type singed gene product is needed for the proper formation of bristles. It is also needed for the production of healthy, fertile eggs. We know this fact because females that are homozygous for certain singed mutations are completely sterile; they lay flimsy, ill-formed eggs that never hatch. However, these mutations have no adverse effect on male fertility. Thus, the singed gene pleiotropically controls the formation of both bristles and eggs in females and the formation of bristles in males.






